- 移动端


上海邦景实业有限公司
9 年
手机商铺
商家活跃:
产品热度:
- NaN
- 0
- 0
- 2
- 2
全部产品
- 查看全部分类
- 菌种
- ATCC细胞
- elisa检测试剂盒
- 抗体
- 标准品
- 培养基
- 试剂盒
- 生化试剂
- elisa酶联免疫试剂盒
- 科研标准品
- 进口ELISA Kit
- RNA/DNA提取
- Sigma试剂
- 科研抗体
- 标准品、对照品
- 代理品牌
- 生物碱
- 黄酮
- 药物杂质及中间体
- 其它天然产物
- 蒽醌
- 其它醌类
- 甾体
- 查尔酮
- 氧杂蒽酮
- 木脂素
- 其它萜类
- 环烯醚萜
- 倍半萜
- 二萜
- 耗材仪器
- 其它酚类
- 三萜
- 香豆素
- 苯丙素
- 中药对照品
- 检测试剂盒
- PCR检测试剂盒
- 生物学试剂
- 细胞库
- 分析化学
- PCR试剂盒
- 耗材和仪器
- PCR鉴定试剂盒
- 进口抗体
- 原代细胞
- LAMP试剂盒
- 滴定缓冲溶液
- 植物提取对照品
- ELISA Kit
- 进口PCR试剂盒
- 进口ELISA检测试剂盒
- 抑制剂
- 快速检测试剂盒
- 荧光定量染料法PCR试剂盒
- 分子化合物
- PCR相关
- RNA相关产品
- 其他DNA/RNA聚合酶
- 核酸纯化
- PCR及RT-PCR相关
- DNA分子量标准
- 克隆载体及相关产品
- 恒温扩增系列
- 蛋白相关
- 核酸酶系列
- 核酸修饰酶系列
- miRNA检测系列
- 提取试剂盒
- 核酸纯化专题
- ELISA试剂盒
- 细胞系
- 蛋白
多功能DNA修复酶抗体
询价
推荐产品
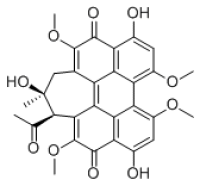
公司新闻/正文
这些进展也提供了一个强有力的、新的、用来阐明Yvc1 功能域和可能的上游信号通路调控的实验系统
405 人阅读发布时间:2018-09-21 10:01
1 Mid1和Cch1
在质膜上, 高亲和力Ca2+内流是由与哺乳动物 Ca2+通道门控电压α1亚单位同源的Cch1和一个可能的门控激活Ca2+通道蛋白Mid1组成的复合体介导的。这个通道与信息交配素存在、内质网压力和内质网/分泌路径储存Ca2+的排空等条件下Ca2+的流入相关。钙调磷酸酶可能通过直接让通道蛋白去磷酸化而抑制通道活性。有至少一个其他的Ca2+内流途径没有得到鉴定, 它可能在交配过程后期介导低亲和力Ca2+内流和细胞融合。
Cch1是酵母中的动物VGCCs(压力门控钙通道)同源物, 它的催化亚基含有四个重复的核心跨膜域。缺少定位于高尔基体上的Ca2+泵-Pmr1的突变株, 显示出很高的依赖于Cch1功能的Ca2+内流比率, 以及高浓度的细胞质自由Ca2+。细胞过表达Pmc1 或者Vcx1时, 也显示依赖Cch1的Ca2+高内流比率, 因为它们与Pmrl竞争底物。此外, Mid1在这些条件下也是Ca2+内流所必需的。在体内, 很多条件下Mid1与 Cch1结合, 也是Cch1产生活性所必需的。这说明Mid1作为Cch1的调节亚单位起作用, 这与动物 VGCCs的α2-Δ亚单位类似。
2 Yvc1
酵母细胞中, 作为Ca2+主要储存器的液泡, 是类似于动物溶酶体的大容量细胞器。与哺乳动物瞬时感受器电位(TRP)通道蛋白同源的酵母Yvc1, 位于液泡膜上。在应答高渗休克和抗心律失常药物胺碘酮时, 从液泡释放Ca2+到细胞质中。然而, 酵母细胞的肌醇1,4,5-三磷酸肌醇(IP3)受体通道还没有被鉴定, Ca2+如何从ER/高尔基体储存中释放出来的问题仍没有研究清楚。
液泡膜的电生理学研究显示, Yvc1对Na+ 、K+ 和Ca2+的选择性透过是完全必需的。在电源性 H+ 泵Vma和次级转运蛋白, 比如H+ -Na+ 交换器(exchanger)Nhx1、H+ -Ca2+交换器Vcx1和门控Cl–通道Gef1的作用下, 液泡包含大量这些阳离子, 细胞质里面微克分子浓度的Ca2+的刺激可以使Yvc1通道打开。另外, Yvc1通道打开的可能性在–80 mV时最大, 当电位更高或者更低时它打开的可能性降低, 这说明它的活性可能受电压变化控制。从这些发现, 可以推断酵母中Yvc1作为一个阳离子释放通道, 起到一个连接到离子库–液泡的作用, 这比胞外环境的离子库更可靠。
Yvc1在体内是怎样受到调控的呢?对Ycv1氨基酸序列的检测没有显示有电压传感蛋白片段, 也没有发现它的核心跨膜域外有与其它蛋白相似的氨基酸序列。然而, 精密的实验揭示, 在活的酵母细胞中, Yvc1在数秒的高渗刺激下被激活。利用细胞质水母荧光素作为细胞溶质自由Ca2+的探针显示, 培养基中高浓度盐或其它渗透物的加入对于细胞质自由CaCa2+浓度的提升, 在Yvc1过表达时得到加强, 在 Yvc1基因敲除时消失。Yvc1-依赖性Ca2+瞬变(transient)显然不需要胞外Ca2+, 但是确实需要Pmc1或者 Vcx1的功能。这些发现说明, Yvc1直接(或者可能间接)引发Ca2+从液泡中释放出来。Yvc1可能也释放其他阳离子, 比如: Na+ 、K+ 和H+ 。这些阳离子被认为在高渗冲击时能够给细胞质提供渗透物以及短暂缓解细胞的脱水状态。Ca2+的释放也能帮助激活本身具有提高高盐抗性的钙调蛋白(calmodulin)和钙调磷酸酶(calcineurin)。这样, 在应对高渗刺激的反应中, Yvc1离子通道处在起中枢作用的位置。这些进展也提供了一个强有力的、新的、用来阐明Yvc1 功能域和可能的上游信号通路调控的实验系统。
对酵母细胞中Ca2+释放调控的观察迫切需要提出这样一个问题: 酵母细胞Ca2+的释放是否伴随着Ca2+的内流呢?在大多数哺乳动物细胞中, Ca2+从内质网中的释放导致质膜上Ca2+内流通道被激活, 是通过被称作储存调控或者容量钙离子进入(CCE)的机制完成的。目前, 对CCE分子水平的了解还很少, 对这个课题的研究可以从像酵母这样的遗传系统获益。酵母液泡不能发起CCE样的过程, 因为缺少 Pmc1和Vcx1的突变株快速地排出液泡内的Ca2+, 并且这些突变株也显示了与野生型相同的通过质膜的Ca2+内流比率。在酵母细胞中, 内质网、高尔基体排出Ca2+或者两者都排出 Ca2+, 确实显现出Ca2+内流的激活。非常有意思的是, 这种由Ca2+排出导致的Ca2+内流需要Cch1。
在质膜上, 高亲和力Ca2+内流是由与哺乳动物 Ca2+通道门控电压α1亚单位同源的Cch1和一个可能的门控激活Ca2+通道蛋白Mid1组成的复合体介导的。这个通道与信息交配素存在、内质网压力和内质网/分泌路径储存Ca2+的排空等条件下Ca2+的流入相关。钙调磷酸酶可能通过直接让通道蛋白去磷酸化而抑制通道活性。有至少一个其他的Ca2+内流途径没有得到鉴定, 它可能在交配过程后期介导低亲和力Ca2+内流和细胞融合。
Cch1是酵母中的动物VGCCs(压力门控钙通道)同源物, 它的催化亚基含有四个重复的核心跨膜域。缺少定位于高尔基体上的Ca2+泵-Pmr1的突变株, 显示出很高的依赖于Cch1功能的Ca2+内流比率, 以及高浓度的细胞质自由Ca2+。细胞过表达Pmc1 或者Vcx1时, 也显示依赖Cch1的Ca2+高内流比率, 因为它们与Pmrl竞争底物。此外, Mid1在这些条件下也是Ca2+内流所必需的。在体内, 很多条件下Mid1与 Cch1结合, 也是Cch1产生活性所必需的。这说明Mid1作为Cch1的调节亚单位起作用, 这与动物 VGCCs的α2-Δ亚单位类似。
2 Yvc1
酵母细胞中, 作为Ca2+主要储存器的液泡, 是类似于动物溶酶体的大容量细胞器。与哺乳动物瞬时感受器电位(TRP)通道蛋白同源的酵母Yvc1, 位于液泡膜上。在应答高渗休克和抗心律失常药物胺碘酮时, 从液泡释放Ca2+到细胞质中。然而, 酵母细胞的肌醇1,4,5-三磷酸肌醇(IP3)受体通道还没有被鉴定, Ca2+如何从ER/高尔基体储存中释放出来的问题仍没有研究清楚。
液泡膜的电生理学研究显示, Yvc1对Na+ 、K+ 和Ca2+的选择性透过是完全必需的。在电源性 H+ 泵Vma和次级转运蛋白, 比如H+ -Na+ 交换器(exchanger)Nhx1、H+ -Ca2+交换器Vcx1和门控Cl–通道Gef1的作用下, 液泡包含大量这些阳离子, 细胞质里面微克分子浓度的Ca2+的刺激可以使Yvc1通道打开。另外, Yvc1通道打开的可能性在–80 mV时最大, 当电位更高或者更低时它打开的可能性降低, 这说明它的活性可能受电压变化控制。从这些发现, 可以推断酵母中Yvc1作为一个阳离子释放通道, 起到一个连接到离子库–液泡的作用, 这比胞外环境的离子库更可靠。
Yvc1在体内是怎样受到调控的呢?对Ycv1氨基酸序列的检测没有显示有电压传感蛋白片段, 也没有发现它的核心跨膜域外有与其它蛋白相似的氨基酸序列。然而, 精密的实验揭示, 在活的酵母细胞中, Yvc1在数秒的高渗刺激下被激活。利用细胞质水母荧光素作为细胞溶质自由Ca2+的探针显示, 培养基中高浓度盐或其它渗透物的加入对于细胞质自由CaCa2+浓度的提升, 在Yvc1过表达时得到加强, 在 Yvc1基因敲除时消失。Yvc1-依赖性Ca2+瞬变(transient)显然不需要胞外Ca2+, 但是确实需要Pmc1或者 Vcx1的功能。这些发现说明, Yvc1直接(或者可能间接)引发Ca2+从液泡中释放出来。Yvc1可能也释放其他阳离子, 比如: Na+ 、K+ 和H+ 。这些阳离子被认为在高渗冲击时能够给细胞质提供渗透物以及短暂缓解细胞的脱水状态。Ca2+的释放也能帮助激活本身具有提高高盐抗性的钙调蛋白(calmodulin)和钙调磷酸酶(calcineurin)。这样, 在应对高渗刺激的反应中, Yvc1离子通道处在起中枢作用的位置。这些进展也提供了一个强有力的、新的、用来阐明Yvc1 功能域和可能的上游信号通路调控的实验系统。
对酵母细胞中Ca2+释放调控的观察迫切需要提出这样一个问题: 酵母细胞Ca2+的释放是否伴随着Ca2+的内流呢?在大多数哺乳动物细胞中, Ca2+从内质网中的释放导致质膜上Ca2+内流通道被激活, 是通过被称作储存调控或者容量钙离子进入(CCE)的机制完成的。目前, 对CCE分子水平的了解还很少, 对这个课题的研究可以从像酵母这样的遗传系统获益。酵母液泡不能发起CCE样的过程, 因为缺少 Pmc1和Vcx1的突变株快速地排出液泡内的Ca2+, 并且这些突变株也显示了与野生型相同的通过质膜的Ca2+内流比率。在酵母细胞中, 内质网、高尔基体排出Ca2+或者两者都排出 Ca2+, 确实显现出Ca2+内流的激活。非常有意思的是, 这种由Ca2+排出导致的Ca2+内流需要Cch1。